








Best Sellers




Featured Deals

Specialty Bars
When the average bar just isn't enough

elitefts Squat Box
EliteFTS Squat Box: Perfect And Take Your Squat To The Next Level

elitefts Patriot Wear
Live Free. Dress Proud.


















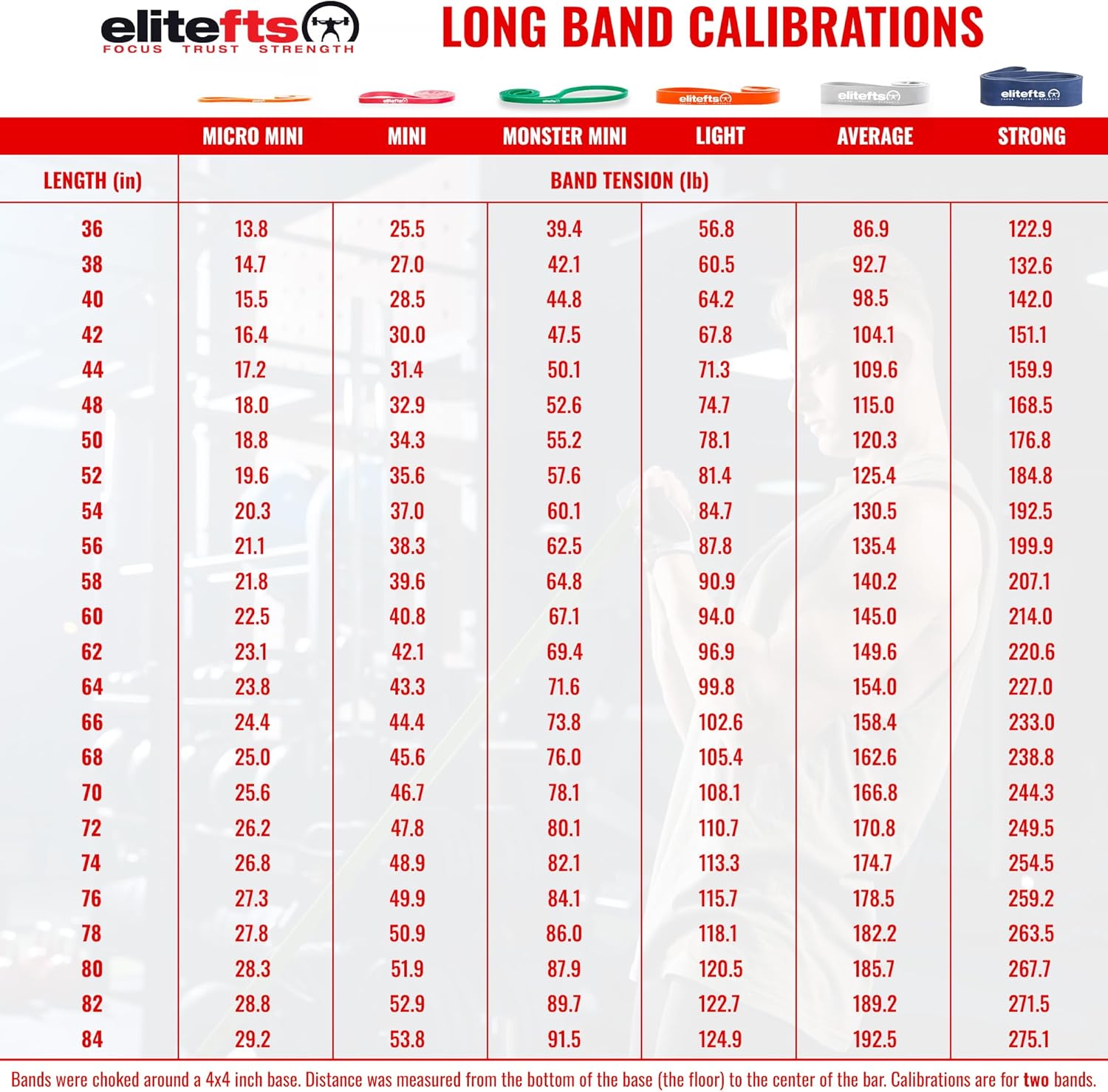


























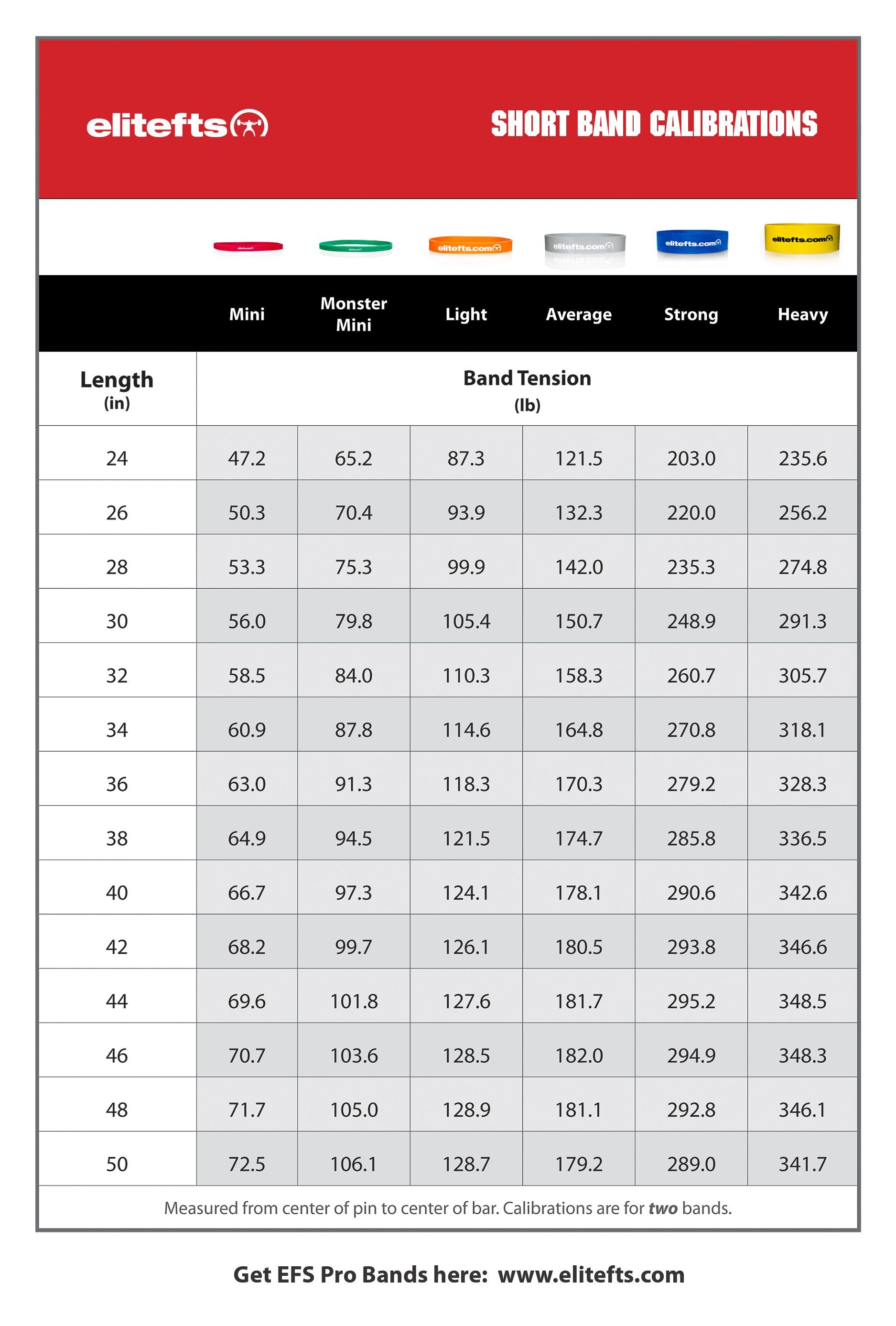







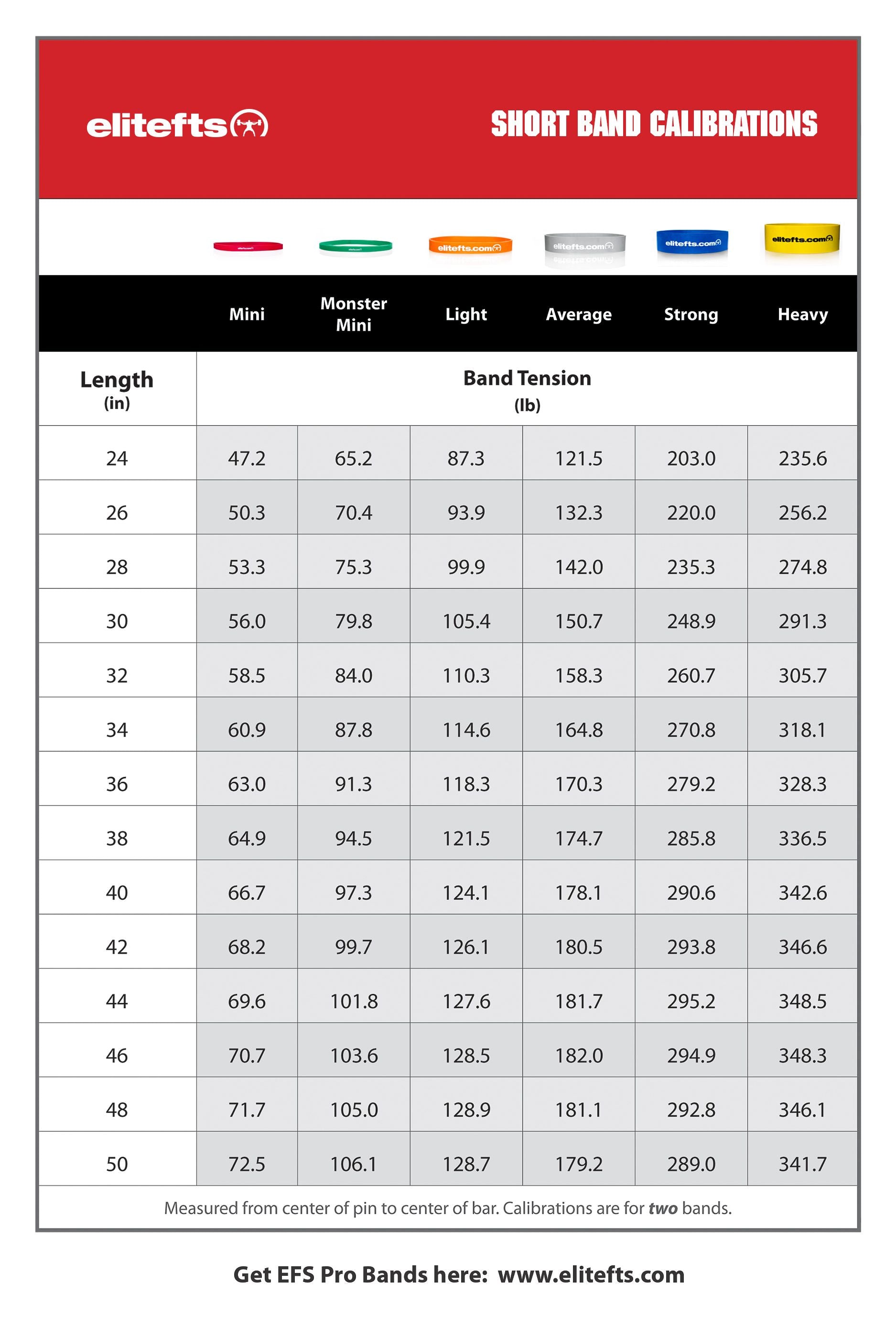











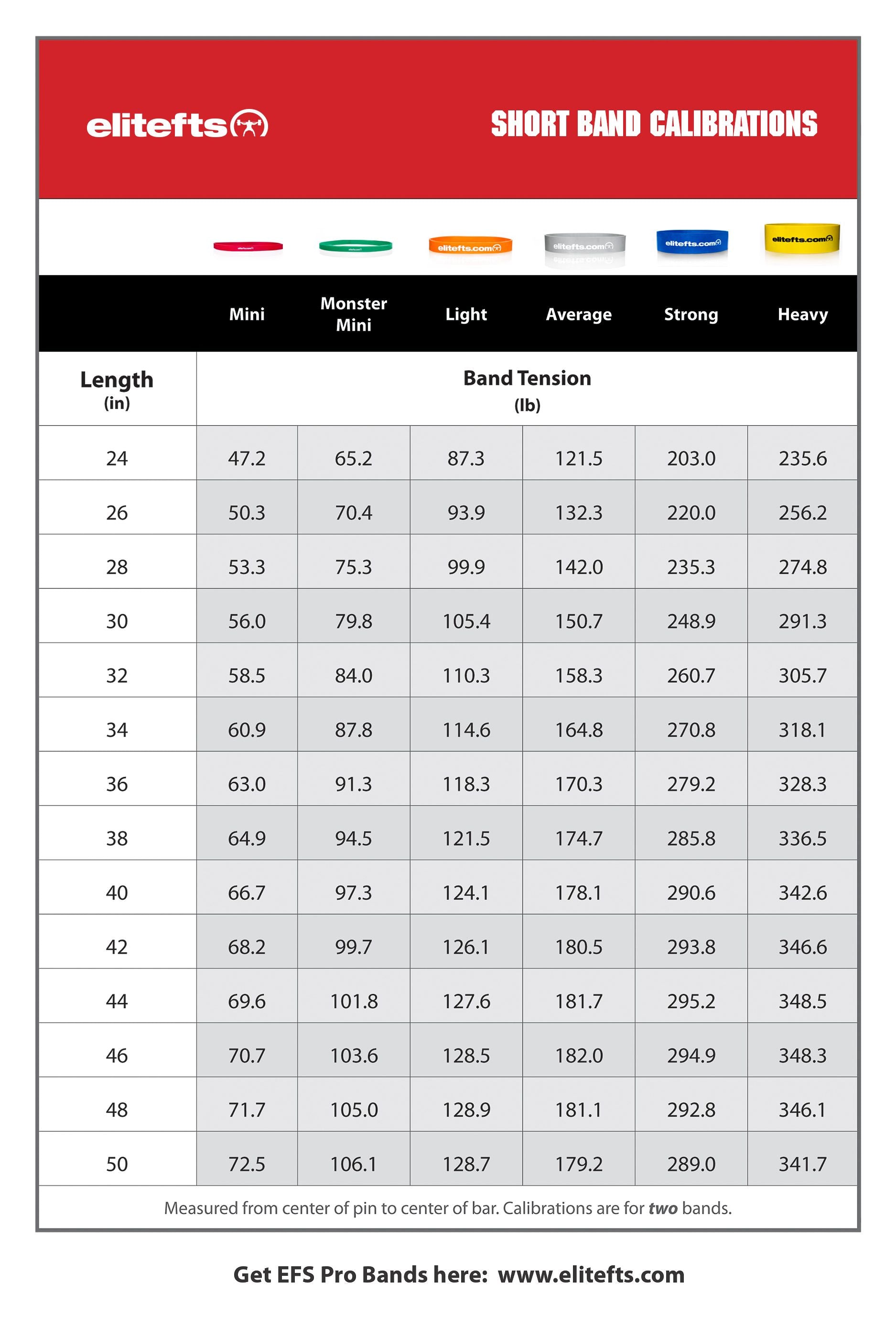




















Education
Trench Built Strength & Conditioning, Workouts, Programs, Training, Coaching and more.

Join The Crew
Support us and access premium content monthly!

Equipment Specialists
Looking for PR-rated custom-built equipment for your home, gym, or school? We have Strength Equipment Specialists to help you out.

Our Company Story
For over 25 years, we’ve remained committed to fostering a culture of excellence, innovation, and integrity.
Blog posts

The Progressive Range of Motion Method: How to Break Your Ceiling Without Adding a Pound to the Bar
A training method built on Paul Anderson's original overload principles that lets advanced lifters break through strength plateaus by systematically raising neurological force output.

Precision Biomechanics for Maximum Back and Chest Hypertrophy
Build a bigger chest and back by learning how elbow path, grip angle, wrist freedom, and joint position determine where tension actually goes. This article breaks down smarter pressing, dips, pulldowns, rows, and EliteFTS gear choices for more precise hypertrophy training.

From Basement Clinics to Better Data: Mike Stratton’s Long Road to Marek Health
From teenage lifter to burned-out founder, Mike Stratton’s story is a raw look at strength culture, hormone health, entrepreneurial sacrifice, and the wake-up call that reshaped his mission with Marek Health.